The gradual stabilization of the NHOS in hepatocytes correlates with their gradual loss of proliferating potential. Nevertheless, the trend towards stabilization of the NHOS with age is observed in both hepatocytes and post-mitotic neurons despite the fact that neurons already have a very stable NHOS from early post-natal age. This fact strongly suggests that thermodynamic and structural constraints drive the NHOS towards maximum stability in time, as the DNA proceeds to dissipate any residual structural stress independently of any cellular functional need. Moreover, considering that entropy is not a measure of disorder or chaos, but of energy diffusion, dissipation or dispersion in a final state compared to an initial state, a rather even distribution of short DNA loops anchored to the NM satisfies the second law of thermodynamics, since the structural stress along the DNA molecule is more evenly dispersed within the nuclear volume by increasing the number of stable DNA-NM interactions. The increase in the relative proportion of total DNA actually embedded within the NM is Lomitapide Mesylate evidence that DNA-NM interactions augment in time leading to further stabilization of the NHOS. The fact that several genes located on different chromosome territories within the Mepiroxol neuronal nucleus lose in time their original privileged position close to the NM, becoming rather distal to it and so they achieve a distribution relative to the NM typical for most random-sequence DNA tracts, indicates that stabilization of the NHOS occurs as a relentless, physical process above any biological constraint. The Fig. 7 depicts two models that may explain how the genes formerly close to the NM become distal to it as a function of time. In the first model genes become distal to the NM because new MARs located far from the genes become actualized substituting in the NM the previous MARs that were close to the genes. Although something like this may be occurring in the case of hepatocytes it is unlikely for the case of neurons considering that there is no real significant change in the already short average DNA-loop size in neurons as a function of age. However the biochemical data indicate that in P540 neurons a significantly larger fraction of total DNA is embedded in the NM when compared with the corresponding fraction in neurons from earlier ages. This fact is consistent with the second model in Fig. 7 which suggests that the actual length of each established MAR increases as a function of time because more DNA ![]() adjacent to the original MAR becomes directly bound to the NM. This results in an overall lesser number of DNA loops with the corresponding displacement of genes to regions relatively distal from the anchoring points to the NM, without significantly changing the average DNA loop size. This model is consistent with our data and with evidence for the existence of rather lengthy MARs, but we cannot directly measure any comparative change in the average MAR length as a function of time since for that purpose we need to destroy the NM in order to liberate the bound MARs, a procedure that leads to unavoidable fragmentation of the formerly bound MARs. Yet, the quantitative changes in the composition of the neuronal NM as a function of age parallel those previously described in aged hepatocytes and may explain how the conditions that allow to increase the direct interactions between DNA and the NM appear in time. Early death is observed in neurons forced to re-enter the cell cycle and neuronal cell cycle activity has been observed early in several diseases that course with neurodegeneration.
adjacent to the original MAR becomes directly bound to the NM. This results in an overall lesser number of DNA loops with the corresponding displacement of genes to regions relatively distal from the anchoring points to the NM, without significantly changing the average DNA loop size. This model is consistent with our data and with evidence for the existence of rather lengthy MARs, but we cannot directly measure any comparative change in the average MAR length as a function of time since for that purpose we need to destroy the NM in order to liberate the bound MARs, a procedure that leads to unavoidable fragmentation of the formerly bound MARs. Yet, the quantitative changes in the composition of the neuronal NM as a function of age parallel those previously described in aged hepatocytes and may explain how the conditions that allow to increase the direct interactions between DNA and the NM appear in time. Early death is observed in neurons forced to re-enter the cell cycle and neuronal cell cycle activity has been observed early in several diseases that course with neurodegeneration.
Reactivated post-mitotic TD cells such as myotubes die very quickly from apotosis
Leave a reply
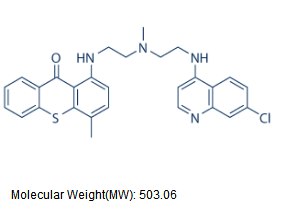 the cell proliferating potential and progression towards terminal differentiation of the hepatocytes with age. We have also compared the NHOS of aged rat hepatocytes with that of early post-mitotic rat neurons, the results indicated that a very stable NHOS is a common feature of both aged and post-mitotic cells in vivo. In the present work we compared the NHOS in rat neurons from different postnatal ages and our results show that the trend towards further stabilization of the NHOS continues in neurons beyond the fourth post-natal week, when the synapses in the cerebral cortex become indistinguishable for those in the adult rat brain and so the neurons are formally regarded as terminally differentiated. This phenomenon occurs in absence of overt changes in the post-mitotic state and transcriptional activity of neurons, suggesting that the continued stabilization of the NHOS as a function of time is basically determined by thermodynamic and structural constraints. We discuss how the resulting highly stable NHOS of neurons may be the non-genetic basis of their permanent and irreversible post-mitotic state. The rat cerebral cortex contains both neurons and glial cells. The classical method by Thompson designed for the isolation of neuronal nuclei separates two nuclear populations: N1 mostly constituted by neuronal nuclei according to old morphological criteria and N2 relatively enriched for glial-cell nuclei on morphological grounds. NeuN/Fox-3 is a protein specific of neuronal nuclei and so labelling with anti-NeuN mAb is the current standard for reliable identification of neurons and neuronal nuclei. In newborn rats the glial cells constitute a small percentage of the brain cells hence there is no point in separating N1 and N2 populations.
the cell proliferating potential and progression towards terminal differentiation of the hepatocytes with age. We have also compared the NHOS of aged rat hepatocytes with that of early post-mitotic rat neurons, the results indicated that a very stable NHOS is a common feature of both aged and post-mitotic cells in vivo. In the present work we compared the NHOS in rat neurons from different postnatal ages and our results show that the trend towards further stabilization of the NHOS continues in neurons beyond the fourth post-natal week, when the synapses in the cerebral cortex become indistinguishable for those in the adult rat brain and so the neurons are formally regarded as terminally differentiated. This phenomenon occurs in absence of overt changes in the post-mitotic state and transcriptional activity of neurons, suggesting that the continued stabilization of the NHOS as a function of time is basically determined by thermodynamic and structural constraints. We discuss how the resulting highly stable NHOS of neurons may be the non-genetic basis of their permanent and irreversible post-mitotic state. The rat cerebral cortex contains both neurons and glial cells. The classical method by Thompson designed for the isolation of neuronal nuclei separates two nuclear populations: N1 mostly constituted by neuronal nuclei according to old morphological criteria and N2 relatively enriched for glial-cell nuclei on morphological grounds. NeuN/Fox-3 is a protein specific of neuronal nuclei and so labelling with anti-NeuN mAb is the current standard for reliable identification of neurons and neuronal nuclei. In newborn rats the glial cells constitute a small percentage of the brain cells hence there is no point in separating N1 and N2 populations.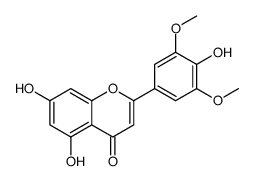 could account for reductions in body/tissue mass accumulation, it has been reported previously that following 90% pancreatectomy the digestive function of the pancreas is well maintained. Moreover, the reduction in body mass observed in our Px animals is consistent with the,20% reduction in mass observed in hyperglycemic Ins2Akita+/2 mice. We also found that direct leucine gavage resulted in an impaired response in mTOR signaling in Px rats, which suggests that a relative reduction in digestive enzymes had a minimal effect on the impaired muscle growth in these animals. Finally, as pointed out in the above discussion, the observations made in this study may only pertain to young Px rats who are not treated with exogenous insulin or allowed physical activity, unlike what is typically done in the clinical care of young patients with T1DM. As such, the relevance that this study has to humans with T1DM remains to be established. In summary, we found that adolescent T1DM skeletal muscle is severely impaired in its capacity for growth, particularly if this occurs at a time when type II fiber development is taking place. The impaired growth can account for the impairments in force production, as force generation made relative to muscle mass was not different between groups. Unlike the other micro- and macrovascular complications associated with long standing diabetes, these differences in muscle growth and resultant decrements in contractile performance exist early on in the disease process. Our data point to impairments in protein synthesis, at a time when these pathways would normally be
could account for reductions in body/tissue mass accumulation, it has been reported previously that following 90% pancreatectomy the digestive function of the pancreas is well maintained. Moreover, the reduction in body mass observed in our Px animals is consistent with the,20% reduction in mass observed in hyperglycemic Ins2Akita+/2 mice. We also found that direct leucine gavage resulted in an impaired response in mTOR signaling in Px rats, which suggests that a relative reduction in digestive enzymes had a minimal effect on the impaired muscle growth in these animals. Finally, as pointed out in the above discussion, the observations made in this study may only pertain to young Px rats who are not treated with exogenous insulin or allowed physical activity, unlike what is typically done in the clinical care of young patients with T1DM. As such, the relevance that this study has to humans with T1DM remains to be established. In summary, we found that adolescent T1DM skeletal muscle is severely impaired in its capacity for growth, particularly if this occurs at a time when type II fiber development is taking place. The impaired growth can account for the impairments in force production, as force generation made relative to muscle mass was not different between groups. Unlike the other micro- and macrovascular complications associated with long standing diabetes, these differences in muscle growth and resultant decrements in contractile performance exist early on in the disease process. Our data point to impairments in protein synthesis, at a time when these pathways would normally be 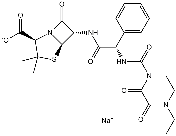 ANK3 is a member of the ankyrin protein family and links integral membrane proteins to the spectrin-based cytoskeleton. It is thought to play a role in the polarized distribution of proteins to specific subcellular sites. ANXA9 belongs to the annexins, a family of evolutionary conserved proteins characterized by their ability to interact with membrane phospholipids in a Ca ++ -dependent way. These proteins have been implicated in membrane organization, membrane-cytoskeleton contacts and vesicular transport. A unique feature of ANXA9 is that its Ca +binding sites are dysfunctional and accordingly its function is unknown. Interestingly, its promoter binds GATA1, an important transcription factor in SC, the expression of which we monitored in our samples. SCIN is an actin-severing protein reported typically in tissues demonstrating a high secretory activity. In bovine SC it was shown to accumulate within the cytoplasm near the base of the cells in a stage-specific manner suggesting a potential role in the
ANK3 is a member of the ankyrin protein family and links integral membrane proteins to the spectrin-based cytoskeleton. It is thought to play a role in the polarized distribution of proteins to specific subcellular sites. ANXA9 belongs to the annexins, a family of evolutionary conserved proteins characterized by their ability to interact with membrane phospholipids in a Ca ++ -dependent way. These proteins have been implicated in membrane organization, membrane-cytoskeleton contacts and vesicular transport. A unique feature of ANXA9 is that its Ca +binding sites are dysfunctional and accordingly its function is unknown. Interestingly, its promoter binds GATA1, an important transcription factor in SC, the expression of which we monitored in our samples. SCIN is an actin-severing protein reported typically in tissues demonstrating a high secretory activity. In bovine SC it was shown to accumulate within the cytoplasm near the base of the cells in a stage-specific manner suggesting a potential role in the 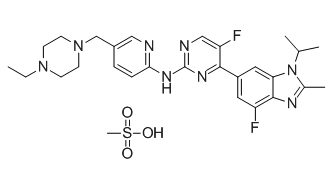 orchestrate the relationship between SC and GC.
orchestrate the relationship between SC and GC.