Thus, low affinity Tulathromycin B interactions might have escaped our detection, but might still have relevance in vivo, if the local concentrations of the interactors are sufficiently high. Notwithstanding the above considerations, a number of controls support the notion that we should have obtained a near complete representation of the EH interactome for EHS-1, ITSN-1 and REPS-1. Conversely, we may have missed a number of interactions for the EH of RME-1, because of the nature of our screening. It has been shown that homo/hetero-oligomerization of EHD proteins is important for optimal binding to NPF-containing proteins, a condition that most likely was not achieved under the conditions of our initial Y2H screening, thus preventing the isolation of strong specific interactors. This is further supported by the fact that the EH domain of RME-1/EHD proteins, located in the carboxylterminal of the proteins, has a strong binding preference for NPF motifs followed by acidic residues. None of the proteins identified in our Y2H screens show an acidic consensus surrounding 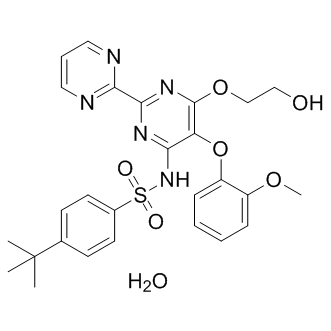 the NPF motif, suggesting that the RME-1 EH binding proteins we identified are probably promiscuous interactors. Indeed, the described interaction between AMPH-1 and RME-1, which was previously shown to be functionally relevant, was not identified in our screening. Regardless of the conditions of screening, it is of note that 14 of the 26 genes encoding for EH-interactors displayed genetic interactions with at least one gene encoding an EH-containing protein. This is remarkable, considering that only one phenotype was analyzed. While a number of these interactions were already known, either in nematodes or in mammals, the others are described here for the first time : together, these interactions define the physical and functional landscape of the EH network at the organismal level in the nematode. As shown in Figure 5, the most evident feature of the EH network is its involvement in LOUREIRIN-B endocytosis, traffic, and actin dynamics. These results confirm the role of the EH network in orchestrating processes in which coordination between the machineries of intracellular traffic and actin remodeling are required. This function is evolutionarily conserved: it has been confirmed in a number of high-resolution studies in mammals, and also by a virtual reconstruction of the EH network in yeast, which we performed by exploiting a number of publicly available interaction data and published high-throughput screens in S. cerevisiae. At the biological level, the EH network seems to play a major role in neurotransmission in the nematode, as supported by the finding that RNAi of the majority of EH interactors affected aldicarb sensitivity either in a WT background or in EH containing proteins mutant strains. While these results can probably be interpreted in the framework of the known participation of EH-containing proteins to the process of synaptic vesicle recycling, through the mentioned connections with endocytosis/traffic and actin dynamics, there is reason to postulate a wider involvement of the EH network in neurotransmission. In particular, the involvement of the EH network in the physiological regulation of the nervous system might also be mirrored by its putative subversion in pathological conditions. Indeed, some of the mammalian homologues of the EHinteracting proteins we identified in the nematode have been implicated in Alzheimer’s disease.
the NPF motif, suggesting that the RME-1 EH binding proteins we identified are probably promiscuous interactors. Indeed, the described interaction between AMPH-1 and RME-1, which was previously shown to be functionally relevant, was not identified in our screening. Regardless of the conditions of screening, it is of note that 14 of the 26 genes encoding for EH-interactors displayed genetic interactions with at least one gene encoding an EH-containing protein. This is remarkable, considering that only one phenotype was analyzed. While a number of these interactions were already known, either in nematodes or in mammals, the others are described here for the first time : together, these interactions define the physical and functional landscape of the EH network at the organismal level in the nematode. As shown in Figure 5, the most evident feature of the EH network is its involvement in LOUREIRIN-B endocytosis, traffic, and actin dynamics. These results confirm the role of the EH network in orchestrating processes in which coordination between the machineries of intracellular traffic and actin remodeling are required. This function is evolutionarily conserved: it has been confirmed in a number of high-resolution studies in mammals, and also by a virtual reconstruction of the EH network in yeast, which we performed by exploiting a number of publicly available interaction data and published high-throughput screens in S. cerevisiae. At the biological level, the EH network seems to play a major role in neurotransmission in the nematode, as supported by the finding that RNAi of the majority of EH interactors affected aldicarb sensitivity either in a WT background or in EH containing proteins mutant strains. While these results can probably be interpreted in the framework of the known participation of EH-containing proteins to the process of synaptic vesicle recycling, through the mentioned connections with endocytosis/traffic and actin dynamics, there is reason to postulate a wider involvement of the EH network in neurotransmission. In particular, the involvement of the EH network in the physiological regulation of the nervous system might also be mirrored by its putative subversion in pathological conditions. Indeed, some of the mammalian homologues of the EHinteracting proteins we identified in the nematode have been implicated in Alzheimer’s disease.
NPF tripeptide at the protein surface or the presence of multiple NPF motifs
Leave a reply