Thus, low affinity Tulathromycin B interactions might have escaped our detection, but might still have relevance in vivo, if the local concentrations of the interactors are sufficiently high. Notwithstanding the above considerations, a number of controls support the notion that we should have obtained a near complete representation of the EH interactome for EHS-1, ITSN-1 and REPS-1. Conversely, we may have missed a number of interactions for the EH of RME-1, because of the nature of our screening. It has been shown that homo/hetero-oligomerization of EHD proteins is important for optimal binding to NPF-containing proteins, a condition that most likely was not achieved under the conditions of our initial Y2H screening, thus preventing the isolation of strong specific interactors. This is further supported by the fact that the EH domain of RME-1/EHD proteins, located in the carboxylterminal of the proteins, has a strong binding preference for NPF motifs followed by acidic residues. None of the proteins identified in our Y2H screens show an acidic consensus surrounding 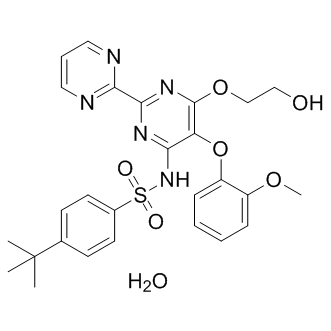 the NPF motif, suggesting that the RME-1 EH binding proteins we identified are probably promiscuous interactors. Indeed, the described interaction between AMPH-1 and RME-1, which was previously shown to be functionally relevant, was not identified in our screening. Regardless of the conditions of screening, it is of note that 14 of the 26 genes encoding for EH-interactors displayed genetic interactions with at least one gene encoding an EH-containing protein. This is remarkable, considering that only one phenotype was analyzed. While a number of these interactions were already known, either in nematodes or in mammals, the others are described here for the first time : together, these interactions define the physical and functional landscape of the EH network at the organismal level in the nematode. As shown in Figure 5, the most evident feature of the EH network is its involvement in LOUREIRIN-B endocytosis, traffic, and actin dynamics. These results confirm the role of the EH network in orchestrating processes in which coordination between the machineries of intracellular traffic and actin remodeling are required. This function is evolutionarily conserved: it has been confirmed in a number of high-resolution studies in mammals, and also by a virtual reconstruction of the EH network in yeast, which we performed by exploiting a number of publicly available interaction data and published high-throughput screens in S. cerevisiae. At the biological level, the EH network seems to play a major role in neurotransmission in the nematode, as supported by the finding that RNAi of the majority of EH interactors affected aldicarb sensitivity either in a WT background or in EH containing proteins mutant strains. While these results can probably be interpreted in the framework of the known participation of EH-containing proteins to the process of synaptic vesicle recycling, through the mentioned connections with endocytosis/traffic and actin dynamics, there is reason to postulate a wider involvement of the EH network in neurotransmission. In particular, the involvement of the EH network in the physiological regulation of the nervous system might also be mirrored by its putative subversion in pathological conditions. Indeed, some of the mammalian homologues of the EHinteracting proteins we identified in the nematode have been implicated in Alzheimer’s disease.
the NPF motif, suggesting that the RME-1 EH binding proteins we identified are probably promiscuous interactors. Indeed, the described interaction between AMPH-1 and RME-1, which was previously shown to be functionally relevant, was not identified in our screening. Regardless of the conditions of screening, it is of note that 14 of the 26 genes encoding for EH-interactors displayed genetic interactions with at least one gene encoding an EH-containing protein. This is remarkable, considering that only one phenotype was analyzed. While a number of these interactions were already known, either in nematodes or in mammals, the others are described here for the first time : together, these interactions define the physical and functional landscape of the EH network at the organismal level in the nematode. As shown in Figure 5, the most evident feature of the EH network is its involvement in LOUREIRIN-B endocytosis, traffic, and actin dynamics. These results confirm the role of the EH network in orchestrating processes in which coordination between the machineries of intracellular traffic and actin remodeling are required. This function is evolutionarily conserved: it has been confirmed in a number of high-resolution studies in mammals, and also by a virtual reconstruction of the EH network in yeast, which we performed by exploiting a number of publicly available interaction data and published high-throughput screens in S. cerevisiae. At the biological level, the EH network seems to play a major role in neurotransmission in the nematode, as supported by the finding that RNAi of the majority of EH interactors affected aldicarb sensitivity either in a WT background or in EH containing proteins mutant strains. While these results can probably be interpreted in the framework of the known participation of EH-containing proteins to the process of synaptic vesicle recycling, through the mentioned connections with endocytosis/traffic and actin dynamics, there is reason to postulate a wider involvement of the EH network in neurotransmission. In particular, the involvement of the EH network in the physiological regulation of the nervous system might also be mirrored by its putative subversion in pathological conditions. Indeed, some of the mammalian homologues of the EHinteracting proteins we identified in the nematode have been implicated in Alzheimer’s disease.
NPF tripeptide at the protein surface or the presence of multiple NPF motifs
Leave a reply
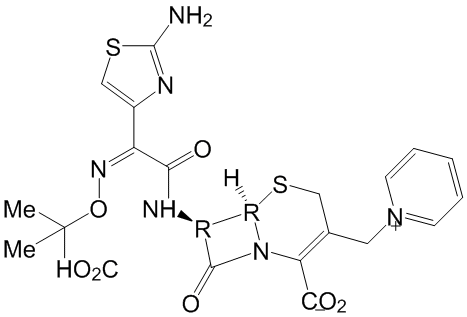 Figure 3A. The tm2156 mutant allele has a deletion of 779 bases resulting in loss of the third intron and of a portion of the fourth exon. reps-1 animals appear to be wild type at different temperatures, in terms of viability, fertility and locomotion. To gain insight into reps-1 functions, we analyzed its expression pattern using transgenic lines carrying the reps-1 gene under its own promoter, in fusion with a GFP reporter. The expression of the fusion protein was analyzed in lysates of transgenic worms by western blot analysis, revealing a protein band with an apparent molecular weight of 75 kDa, in agreement with the predicted molecular weight for REPS-1::GFP. The transgenic lines showed expression in many tissues including intestine, secretory system, vulval cells and muscle cells. REPS-1 was also expressed in the nervous system with diffuse staining in the nerve ring, ventral cord and commissures, but no expression was observed in the neuronal body. When tested for sensitivity to aldicarb, an inhibitor of acetylcholine esterase often used to reveal defective cholinergic transmission, the reps-1 mutant showed an abnormal response, with hypersensitivity to the drug
Figure 3A. The tm2156 mutant allele has a deletion of 779 bases resulting in loss of the third intron and of a portion of the fourth exon. reps-1 animals appear to be wild type at different temperatures, in terms of viability, fertility and locomotion. To gain insight into reps-1 functions, we analyzed its expression pattern using transgenic lines carrying the reps-1 gene under its own promoter, in fusion with a GFP reporter. The expression of the fusion protein was analyzed in lysates of transgenic worms by western blot analysis, revealing a protein band with an apparent molecular weight of 75 kDa, in agreement with the predicted molecular weight for REPS-1::GFP. The transgenic lines showed expression in many tissues including intestine, secretory system, vulval cells and muscle cells. REPS-1 was also expressed in the nervous system with diffuse staining in the nerve ring, ventral cord and commissures, but no expression was observed in the neuronal body. When tested for sensitivity to aldicarb, an inhibitor of acetylcholine esterase often used to reveal defective cholinergic transmission, the reps-1 mutant showed an abnormal response, with hypersensitivity to the drug  force for cellular contractility and lead to the retraction of the trailing edge. Rho small GTPases are clearly pivotal in all of these cytoskeletal rearrangement processes. For example, Rac is primarily responsible for generating a protrusive force through the localized actin polymerization, while Rho is responsible for the
force for cellular contractility and lead to the retraction of the trailing edge. Rho small GTPases are clearly pivotal in all of these cytoskeletal rearrangement processes. For example, Rac is primarily responsible for generating a protrusive force through the localized actin polymerization, while Rho is responsible for the  conjugated to a protein antigen such as bovine serum albumin, significantly increased protection against a type A strain. This highlights the need to identify a diverse set of FT-specific immunodominant antigens that can stimulate antibody and cell-mediated responses to combat FT infection.
conjugated to a protein antigen such as bovine serum albumin, significantly increased protection against a type A strain. This highlights the need to identify a diverse set of FT-specific immunodominant antigens that can stimulate antibody and cell-mediated responses to combat FT infection. respiratory tularemia. Tul4 is a TLR2 agonist. Therefore, in addition to the adjuvant effects of GPI, it is possible that Tul4 might also act in potentiating innate and/or adaptive immune responses. In this regard, it has been shown that innate immune responses induced via several TLRs can protect against a lethal FT challenge : however, for optimal protection, TLR agonists need to be administered 1�C3 days before infection. Furthermore, since TLRs influence adaptive immune responses; it is possible that Tul4mediated TLR signaling might provide additional priming for the development of an antigen-specific response. This possibility gains support from the evidence that Tul4 induces a robust antibody response even in the absence of an adjuvant, and from previous studies demonstrating the importance of TLR2 in host responses to F. tularensis. Cytokines are important players of immune responses, and several cytokines, including TNF-a, IFN-c, IL-6, IL-12, IL-17A and IL-10, have been shown to be relevant in the host defense against FT infection. However, our studies did not reveal a specific cytokine pattern associated with protection seen in DnaK+Tul4+GPI immunized mice. In this regard, no difference was observed in the level or pattern of the IFN-c response among all experimental groups of mice, even though IFN-c has been shown to be a critical cytokine in host responses against FT infection. Moreover, between 3�C5 days after infection, the levels of IL-12p70, TNF-a and IL-6 were higher in mice immunized with one of the antigens than in mice immunized with both DnaK and Tul4, a pattern similar to that seen with the other cytokines assessed. These findings suggest that in vivo, protection against FT infection likely occurs due to the presence and interactions of numerous inflammatory mediators. Molecules like IP-10, CXCL1, RANTES and G-CSF that were evaluated in the present study are also critical participants in immune responses. For instance, RANTES, IP-10 and CXCL1, are chemotactic cytokines that belong to the chemokine family and play active roles in recruiting leukocytes to inflammatory sites. Furthermore, G-CSF, besides being a growth factor, is a cytokine that stimulates the survival, proliferation, differentiation and function of neutrophils, in addition to its involvement in the regulation of the PI3K/Akt signaling pathway. Finally, the observed changes in cytokine levels between 3 to 5 days after infection are perhaps important for the observed protection, since a differential modulation of the cytokines during this period was observed between infected mice immunized with each antigen and mice immunized with the
respiratory tularemia. Tul4 is a TLR2 agonist. Therefore, in addition to the adjuvant effects of GPI, it is possible that Tul4 might also act in potentiating innate and/or adaptive immune responses. In this regard, it has been shown that innate immune responses induced via several TLRs can protect against a lethal FT challenge : however, for optimal protection, TLR agonists need to be administered 1�C3 days before infection. Furthermore, since TLRs influence adaptive immune responses; it is possible that Tul4mediated TLR signaling might provide additional priming for the development of an antigen-specific response. This possibility gains support from the evidence that Tul4 induces a robust antibody response even in the absence of an adjuvant, and from previous studies demonstrating the importance of TLR2 in host responses to F. tularensis. Cytokines are important players of immune responses, and several cytokines, including TNF-a, IFN-c, IL-6, IL-12, IL-17A and IL-10, have been shown to be relevant in the host defense against FT infection. However, our studies did not reveal a specific cytokine pattern associated with protection seen in DnaK+Tul4+GPI immunized mice. In this regard, no difference was observed in the level or pattern of the IFN-c response among all experimental groups of mice, even though IFN-c has been shown to be a critical cytokine in host responses against FT infection. Moreover, between 3�C5 days after infection, the levels of IL-12p70, TNF-a and IL-6 were higher in mice immunized with one of the antigens than in mice immunized with both DnaK and Tul4, a pattern similar to that seen with the other cytokines assessed. These findings suggest that in vivo, protection against FT infection likely occurs due to the presence and interactions of numerous inflammatory mediators. Molecules like IP-10, CXCL1, RANTES and G-CSF that were evaluated in the present study are also critical participants in immune responses. For instance, RANTES, IP-10 and CXCL1, are chemotactic cytokines that belong to the chemokine family and play active roles in recruiting leukocytes to inflammatory sites. Furthermore, G-CSF, besides being a growth factor, is a cytokine that stimulates the survival, proliferation, differentiation and function of neutrophils, in addition to its involvement in the regulation of the PI3K/Akt signaling pathway. Finally, the observed changes in cytokine levels between 3 to 5 days after infection are perhaps important for the observed protection, since a differential modulation of the cytokines during this period was observed between infected mice immunized with each antigen and mice immunized with the