Thus, the disease-associated P1018L mutation conferred rapid inactivation of 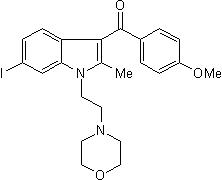 the hTRPM2 channel, whereas manipulation of the pore region by site-directed mutagenesis resulted in a TRPM2-LDE mutant channel that exhibited no inactivation, suggesting alterations in the conformation and structure of the pore region represent an important molecular mechanisms of the TRPM2 channel inactivation. Ion channels play important roles in diverse physiological processes, including neuronal signaling, cardiac rhythm setting, and insulin secretion. Small molecules that positively or negatively modulate ion channel activities are commonly used as drugs or research tools. Open-channel blockers, a class of ion channel inhibitors that sterically occlude the open pore of an active channel, have been used in both clinical treatment and basic research. For instance, slow-channel congenital myasthenic syndromes, a type of neuromuscular junction dysfunctions MDL-29951 caused by abnormally prolonged opening of the nicotinic acetylcholine receptor, are treated by long-lived blockers for the AChR. Memantine, an open-channel blocker for the N-methyl-Daspartate receptor, is used for the treatment of Alzheimer’s disease. Due to the versatility and clinical potential of open-channel blockers, it would be desirable to develop new strategies for more convenient and straightforward design of these agents. Rational design of open-channel blockers has been challenging due to the limited availability of high-resolution structures for the ion-conducting pores. Open-channel blockers are believed to bind within the transmembrane pore via noncovalent interactions with the hydrophobic pore-lining residues. Drugs that exhibit appreciable blockade affinities mostly have rigid and/or complex structures, suggesting that these molecules have well-defined binding geometries within their target sites. Without a detailed structure of the open channel pore, blocker design or optimization cannot be carried out via the typical structure-based approach, in which a ligand needs to be docked in the binding site. Even when a high-resolution structure of the target channel is available, it remains difficult to predict how variations in the chemical structure would affect the kinetics of blocker association and dissociation. AGEs accumulate over time and are used as markers of carbonyl stress. Advanced glycation endproducts are very stable and protease-resistant. Therefore, AGE-induced crosslinks of peptides and proteins lead to protein deposition and amyloidosis, which is a reason why AGEs are involved in a variety of diseases, such as arteriosclerosis, diabetic nephropathy and Etidronate neuropathy or cataract. As an example, AGEs have been detected in vascular walls, glomerular basement membranes and the renal cortex, as well as in amyloid plaques in Alzheimers disease. During AGE formation, oxygen radicals are also generated, which, beside the AGEs themselves, are involved in neuronal cell damage by oxygen stress and apoptotic processes. Taken together, the glycation theory is one explanation for the molecular mechanism of ageing. In line with this theory, crosslinking and denaturation of proteins caused by glycation, are major factors for early ageing. However, glycation of proteins is not the only mechanism of ageing. Oxidative damage by reactive oxygen species is also involved in ageing. This implies that every cell is under constant pressure to remove waste that accumulates in form of metabolically damaged proteins and xenobiotics. Preventing the generating of these metabolic waste products help to prevent cellular ageing and has been termed the ”green theory” of ageing. In this study we focused on the involvement of AGEs on neuronal adhesion and differentiation because age-related accumulation of AGEs.
the hTRPM2 channel, whereas manipulation of the pore region by site-directed mutagenesis resulted in a TRPM2-LDE mutant channel that exhibited no inactivation, suggesting alterations in the conformation and structure of the pore region represent an important molecular mechanisms of the TRPM2 channel inactivation. Ion channels play important roles in diverse physiological processes, including neuronal signaling, cardiac rhythm setting, and insulin secretion. Small molecules that positively or negatively modulate ion channel activities are commonly used as drugs or research tools. Open-channel blockers, a class of ion channel inhibitors that sterically occlude the open pore of an active channel, have been used in both clinical treatment and basic research. For instance, slow-channel congenital myasthenic syndromes, a type of neuromuscular junction dysfunctions MDL-29951 caused by abnormally prolonged opening of the nicotinic acetylcholine receptor, are treated by long-lived blockers for the AChR. Memantine, an open-channel blocker for the N-methyl-Daspartate receptor, is used for the treatment of Alzheimer’s disease. Due to the versatility and clinical potential of open-channel blockers, it would be desirable to develop new strategies for more convenient and straightforward design of these agents. Rational design of open-channel blockers has been challenging due to the limited availability of high-resolution structures for the ion-conducting pores. Open-channel blockers are believed to bind within the transmembrane pore via noncovalent interactions with the hydrophobic pore-lining residues. Drugs that exhibit appreciable blockade affinities mostly have rigid and/or complex structures, suggesting that these molecules have well-defined binding geometries within their target sites. Without a detailed structure of the open channel pore, blocker design or optimization cannot be carried out via the typical structure-based approach, in which a ligand needs to be docked in the binding site. Even when a high-resolution structure of the target channel is available, it remains difficult to predict how variations in the chemical structure would affect the kinetics of blocker association and dissociation. AGEs accumulate over time and are used as markers of carbonyl stress. Advanced glycation endproducts are very stable and protease-resistant. Therefore, AGE-induced crosslinks of peptides and proteins lead to protein deposition and amyloidosis, which is a reason why AGEs are involved in a variety of diseases, such as arteriosclerosis, diabetic nephropathy and Etidronate neuropathy or cataract. As an example, AGEs have been detected in vascular walls, glomerular basement membranes and the renal cortex, as well as in amyloid plaques in Alzheimers disease. During AGE formation, oxygen radicals are also generated, which, beside the AGEs themselves, are involved in neuronal cell damage by oxygen stress and apoptotic processes. Taken together, the glycation theory is one explanation for the molecular mechanism of ageing. In line with this theory, crosslinking and denaturation of proteins caused by glycation, are major factors for early ageing. However, glycation of proteins is not the only mechanism of ageing. Oxidative damage by reactive oxygen species is also involved in ageing. This implies that every cell is under constant pressure to remove waste that accumulates in form of metabolically damaged proteins and xenobiotics. Preventing the generating of these metabolic waste products help to prevent cellular ageing and has been termed the ”green theory” of ageing. In this study we focused on the involvement of AGEs on neuronal adhesion and differentiation because age-related accumulation of AGEs.
Mutation of the residues in the pore region can strongly alter the channel inactivation
Leave a reply