Cryostat sections 10 mm thick were cut at the greatest diameter of tumor, mounted on glass slides, and allowed to air-dry. Doxorubicin auto-fluorescence was detected using Nikon TE2000 inverted fluorescence microscopy 465 to 495 nm excitation and 510 to 555 nm emission filters, and images of tissue section were captured with a Nikon DS-U3 AbMole Corosolic-acid camera. Blood vessels were recognized by the expression of CD31 membrane protein on endothelial cells. Finally, sections were cover-slipped with a mounting medium containing anti-fluorescein quencher. Anti-CD31 fluorescence representing endothelial cell was detected with 510 to 560 nm excitation and 590 nm emission filters, and DAPI-stained nuclear DNA localization was detected with 330�C380 nm excitation and 420 nm emission filters. Composite images of doxorubicin and CD31 fluorescence were generated using Image Pro Plus software. For CD31 images, Microvessel density was evaluated using the counting method introduced by Weidner et al. The tumor sections were firstly scanned at low magnification field to find the areas that showed the most intense vascularization and then individual microvessels were counted in three different high magnification fields. The final MVD was the mean value of the counts of the three fields. For the evaluation of doxorubicin, three different high magnification fields with high doxorubicin fluorescence were selected. Areas of necrosis and staining artifact were excluded. The penetration of doxorubicin was measured using the Image Pro Plus software application and customized algorithms. Briefly, a doxorubicin image and a separate blood vessel image of the same field were firstly captured. In order to make the blood vessel image the same size as the doxorubicin image, an extra copy of the doxorubicin image was made, and was overlaid on top of the blood vessel image. The blood vessel image was then masked, so that all the blood vessels can be identified and be converted to white color with an intensity of 255. The remaining pixels in that image had an intensity of 0. This was called a binarized image. Any pixel that was not 0 was AbMole Mepiroxol regarded as a blood vessel. The image then was converted and a new image was formed by using the function of 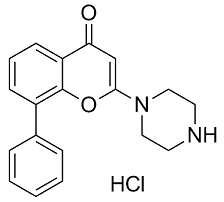 distance filter. In this picture, the gray value of each point represented the distance of the point to the nearest vessel. The doxorubicin image was also switched to the binarized image in which each doxorubicin positive spot was represented as its center point. The data were tabulated to average out the doxorubicin penetration distance of the field. The mean penetration distance of the three fields was taken as the final doxorubicin penetration distance for each animal. The count of doxorubicin-specific fluorescent spot in each field was simultaneously counted.
distance filter. In this picture, the gray value of each point represented the distance of the point to the nearest vessel. The doxorubicin image was also switched to the binarized image in which each doxorubicin positive spot was represented as its center point. The data were tabulated to average out the doxorubicin penetration distance of the field. The mean penetration distance of the three fields was taken as the final doxorubicin penetration distance for each animal. The count of doxorubicin-specific fluorescent spot in each field was simultaneously counted.
It is well known that to be most effective chemotherapeutic agents must penetrate deep
Leave a reply
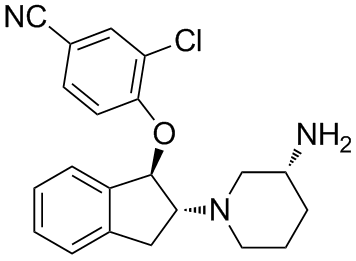 colicin A was also found to resemble Bcl-xL, which belongs to the Bcl-2 family and is a regulator of programmed cell death. This finding notably led to the discovery of the pore-forming activity of Bcl-xL. The other examples are colicin E5 and colicin D, which cleave the anticodon loops of specific transfer RNAs. Colicin E5 recognizes the QpUp sequence and mediates tRNA cleavage between positions 34 and 35 in tRNATyr, tRNAHis, tRNAAsn, and tRNAAsp in sensitive E. coli cells. Similarly, colicin D targets 4 iso-accepting tRNA Args and cleaves between positions 38 and 39. These findings suggest that tRNA cleavage induces DNA damage. However, when D-CRD, an active domain of colicin D, was ectopically expressed in yeast cells with genetic defects in the DNA repair system, none of the cells displayed this altered sensitivity. Although the tRNA species cleaved by D-CRD are different from those cleaved by zyomcin and PaT, this result suggested that DNA damage is not an indirect consequence of tRNA cleavage, and we tried to elucidate the mechanism by which PaT Orf2p induces DNA damage. We then found evidence that PaT Orf2p cleaves DNA, in addition to its specific tRNA cleavage activity. Moreover, the translation impairment caused by tRNA cleavage allows
colicin A was also found to resemble Bcl-xL, which belongs to the Bcl-2 family and is a regulator of programmed cell death. This finding notably led to the discovery of the pore-forming activity of Bcl-xL. The other examples are colicin E5 and colicin D, which cleave the anticodon loops of specific transfer RNAs. Colicin E5 recognizes the QpUp sequence and mediates tRNA cleavage between positions 34 and 35 in tRNATyr, tRNAHis, tRNAAsn, and tRNAAsp in sensitive E. coli cells. Similarly, colicin D targets 4 iso-accepting tRNA Args and cleaves between positions 38 and 39. These findings suggest that tRNA cleavage induces DNA damage. However, when D-CRD, an active domain of colicin D, was ectopically expressed in yeast cells with genetic defects in the DNA repair system, none of the cells displayed this altered sensitivity. Although the tRNA species cleaved by D-CRD are different from those cleaved by zyomcin and PaT, this result suggested that DNA damage is not an indirect consequence of tRNA cleavage, and we tried to elucidate the mechanism by which PaT Orf2p induces DNA damage. We then found evidence that PaT Orf2p cleaves DNA, in addition to its specific tRNA cleavage activity. Moreover, the translation impairment caused by tRNA cleavage allows 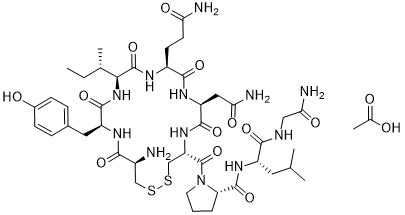 detail. Here we describe a general, simple and rapid approach to monitor the protein binding on an aptamer-based biosensor and independently assess both the aptamer-protein solution- and surface-phase affinities. The main advantages of our approach are the facts that the determination of the solution-phase affinity is done before reaching the equilibrium state on the biosensor, which is generally time consuming especially at low concentrations of proteins and is independent of a surface-phase model of adsorption. In the later case, the Langmuir model is usually considered for simplicity, but the use of more sophisticated models may be required to take into account mutual interactions, heterogeneity and loss of accessibility of the probes. A proof-of-principle label-free kinetic biosensor was developed, using the two advantageous traits of Surface Plasmon Resonance imaging at their best : the label-free and realtime detection of molecular interactions occurring on the biosensor surface. State-of-the-art performances for a label-free technique without amplification were obtained: a sub-nanomolar limit of detection with a linear range of quantification of two orders of magnitude while a rapid detection and a strong selectivity were maintained. The independent determinations of KDSol and KDSurf, respectively the solution- and surface-phase affinities, revealed distinct values illustrating the importance of probes, targets or surface interactions in biosensors. For this purpose, an
detail. Here we describe a general, simple and rapid approach to monitor the protein binding on an aptamer-based biosensor and independently assess both the aptamer-protein solution- and surface-phase affinities. The main advantages of our approach are the facts that the determination of the solution-phase affinity is done before reaching the equilibrium state on the biosensor, which is generally time consuming especially at low concentrations of proteins and is independent of a surface-phase model of adsorption. In the later case, the Langmuir model is usually considered for simplicity, but the use of more sophisticated models may be required to take into account mutual interactions, heterogeneity and loss of accessibility of the probes. A proof-of-principle label-free kinetic biosensor was developed, using the two advantageous traits of Surface Plasmon Resonance imaging at their best : the label-free and realtime detection of molecular interactions occurring on the biosensor surface. State-of-the-art performances for a label-free technique without amplification were obtained: a sub-nanomolar limit of detection with a linear range of quantification of two orders of magnitude while a rapid detection and a strong selectivity were maintained. The independent determinations of KDSol and KDSurf, respectively the solution- and surface-phase affinities, revealed distinct values illustrating the importance of probes, targets or surface interactions in biosensors. For this purpose, an