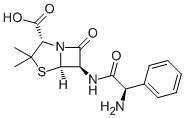
On post-translational activation and stability of cell cycle regulatory proteins. To identify downstream molecular actors involved in PTEN mediated cell cycle modulation, a prostatic cancer cellular model expressing PTEN cDNA was recently used. This modification led to a significant inhibition of cell proliferation due to cycle arrest in the G1 phase. Gene expression analysis subsequent to PTEN reintroduction, coupled with phosphorylation status of downstream molecular targets, suggest that PTEN physiologically regulates cell cycle related proteins, through pAKT dependent and independent ways. E2F2, cdc25a, Cyclin G2 and RBL2 proteins are among them. The aim of the present study was to shed light on the role of PTEN pathway on cell cycle regulation in GSCs through a cellular differentiation model. Phosphoproteomics profiling of different GSCs lines, using reverse phase protein microarrays, indicates a heterogeneous basal activity of PTEN and related molecules. Transcriptomics profiling of GSCs induced to differentiate demonstrates specific molecular changes in PTEN positive cells, that are not observed in cells lacking its activation. The most modulated transcripts suggest a post-transcriptional regulation model, which involves cdc25a as the main target. A diagnosis of glioblastoma multiforme, established histologically according to the WHO classification, was the eligibility criteria. Patients AbMole Metyrapone characteristics are summarized in table 1. Tumor samples were obtained by complete or partial resection before the initiation of treatment with radiation and chemotherapy. The expression of both the proliferation marker Ki-67 and of tumor protein 53 were characterized on tumor specimen by 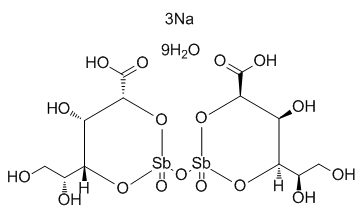 immunohistochemistry on deparaffinized sections using the avidin-biotin-peroxidase complex methods, anti-p53 monoclonal antibody and AbMole Mepiroxol anti-epidermal growth factor receptor monoclonal antibody. Tumors were considered p53 deficient if immunoreaction stained the nuclei of less than 5% of cells. Tumors showing moderate-to-strong immunostaining for EGFR in.20% of cells were considered EGFR positive. O6-methylguanine-DNA methyltransferase promoter methylation patterns were assessed on genomic DNA extracted from paraffin-embedded tissue by methylation-specific: it has been shown that the DNA repair protein MGMT influence the resistance of glioblastoma cells to alkylating agents, such as nitrosoureas and temozolomide thus representing and important prognostic and therapeutic indicator. Stem cells tightly regulate cell cycle progression in order to remain quiescent inside their cellular niche with progress occurring only when external stimuli indicates the need for cell growth or for the replacement of damaged or senescent cells.
immunohistochemistry on deparaffinized sections using the avidin-biotin-peroxidase complex methods, anti-p53 monoclonal antibody and AbMole Mepiroxol anti-epidermal growth factor receptor monoclonal antibody. Tumors were considered p53 deficient if immunoreaction stained the nuclei of less than 5% of cells. Tumors showing moderate-to-strong immunostaining for EGFR in.20% of cells were considered EGFR positive. O6-methylguanine-DNA methyltransferase promoter methylation patterns were assessed on genomic DNA extracted from paraffin-embedded tissue by methylation-specific: it has been shown that the DNA repair protein MGMT influence the resistance of glioblastoma cells to alkylating agents, such as nitrosoureas and temozolomide thus representing and important prognostic and therapeutic indicator. Stem cells tightly regulate cell cycle progression in order to remain quiescent inside their cellular niche with progress occurring only when external stimuli indicates the need for cell growth or for the replacement of damaged or senescent cells.
Investigating the PTEN-dependent cell cycle regulation reported the the influence of PTEN/Akt signaling pathway
Leave a reply
 is optimized for two conflicting objective functions, respectively. In LAT-deficient J.Cam2 and ANJ3 cells, PLCc1, VAV and SLP76 phosphorylation were not detected and TCR-mediated signaling events were found to be impaired. Early T cell development has been found to be blocked in LAT-negative mice similar to that observed in mice, in which either LCK and FYN, or ZAP70 and Syk was deleted. These observations demonstrate that LAT is an obligatory step, downstream of LCK and ZAP70, and is an important link to Ras/ MAP kinase and PLCc1 pathways. Dlg1 is expressed in all T cell developmental stages and forms a stable complex with LCK. It functions as a negative regulator of T cell activation. Endogenous Dlg1 connects with TCRf and CBL. The transient overexpression of Dlg1 attenuates basal and VAV1-induced NFAT reporter activation. It is also shown that the reduction of Dlg1 expression by RNA interference enhances both CD3- and SAg-mediated NFAT activation. Thus it is interpreted that Dlg1 acts as an activation antagonist. This is the reason, we have got higher c-value for Dlgh1 in an unperturbed TCR signaling pathway. In contrast, it has become 0 and 0.46094 in pathogen perturbed system and in the system, where it is optimized for two conflicting objective functions, respectively. The cytoskeleton of eukaryotic cells has a pivotal role in hostpathogen interactions as it is essential for epithelial and endothelial barrier functions. It limits the invasion and dissemination of pathogens in tissues. GTP-binding proteins of the Rho family are regulators of the actin cytoskeleton, and Rho proteins play essential roles in host cell invasion by bacteria. They are the targets for bacterial protein toxins that either inactivate GTPases by ADP-ribosylation or glucosylation, or activate them by deamidation. Specifically, Rho GTPase Cdc42 regulates cytoskeletal changes at the immunological synapse that are critical to T cell activation.
is optimized for two conflicting objective functions, respectively. In LAT-deficient J.Cam2 and ANJ3 cells, PLCc1, VAV and SLP76 phosphorylation were not detected and TCR-mediated signaling events were found to be impaired. Early T cell development has been found to be blocked in LAT-negative mice similar to that observed in mice, in which either LCK and FYN, or ZAP70 and Syk was deleted. These observations demonstrate that LAT is an obligatory step, downstream of LCK and ZAP70, and is an important link to Ras/ MAP kinase and PLCc1 pathways. Dlg1 is expressed in all T cell developmental stages and forms a stable complex with LCK. It functions as a negative regulator of T cell activation. Endogenous Dlg1 connects with TCRf and CBL. The transient overexpression of Dlg1 attenuates basal and VAV1-induced NFAT reporter activation. It is also shown that the reduction of Dlg1 expression by RNA interference enhances both CD3- and SAg-mediated NFAT activation. Thus it is interpreted that Dlg1 acts as an activation antagonist. This is the reason, we have got higher c-value for Dlgh1 in an unperturbed TCR signaling pathway. In contrast, it has become 0 and 0.46094 in pathogen perturbed system and in the system, where it is optimized for two conflicting objective functions, respectively. The cytoskeleton of eukaryotic cells has a pivotal role in hostpathogen interactions as it is essential for epithelial and endothelial barrier functions. It limits the invasion and dissemination of pathogens in tissues. GTP-binding proteins of the Rho family are regulators of the actin cytoskeleton, and Rho proteins play essential roles in host cell invasion by bacteria. They are the targets for bacterial protein toxins that either inactivate GTPases by ADP-ribosylation or glucosylation, or activate them by deamidation. Specifically, Rho GTPase Cdc42 regulates cytoskeletal changes at the immunological synapse that are critical to T cell activation. similarity between the existing experimental observations and the computed c-values. We have found lower c-value for LCK in an un
similarity between the existing experimental observations and the computed c-values. We have found lower c-value for LCK in an un in a perturbed TCR signaling pathway. It has again decreased in an integrated pathway which was optimized for two conflicting objective functions. MALT1 is recruited to the lipid rafts of the immunological synapse following activation of TCR and CD28 coreceptor. This recruitment is dependent on CARMA1. MALT1, Bcl10, and CARMA1 form a trimolecular complex. Expression of a MALT1 deletion mutant has shown to completely block the CD3/CD28 costimulation-induced NF-kB activation. Considering the above experimental evidences for CARMA complex, we have also found lower c-value for CARMA complex in an unperturbed TCR signaling pathway. In contrast, it has become 0.81185 and 0.23229 in pathogen perturbed system and in the system, where it is optimized for two conflicting objective functions, respectively. NCK plays a pivotal role in TCR-induced reorganization of the actin cytoskeleton and the formation of the immunological synapse in T lymphocytes.
in a perturbed TCR signaling pathway. It has again decreased in an integrated pathway which was optimized for two conflicting objective functions. MALT1 is recruited to the lipid rafts of the immunological synapse following activation of TCR and CD28 coreceptor. This recruitment is dependent on CARMA1. MALT1, Bcl10, and CARMA1 form a trimolecular complex. Expression of a MALT1 deletion mutant has shown to completely block the CD3/CD28 costimulation-induced NF-kB activation. Considering the above experimental evidences for CARMA complex, we have also found lower c-value for CARMA complex in an unperturbed TCR signaling pathway. In contrast, it has become 0.81185 and 0.23229 in pathogen perturbed system and in the system, where it is optimized for two conflicting objective functions, respectively. NCK plays a pivotal role in TCR-induced reorganization of the actin cytoskeleton and the formation of the immunological synapse in T lymphocytes.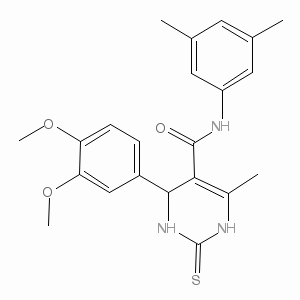 invaginations of the intestinal epithelium; the crypts of Lieberk��hn. The intestinal stem cells are characterized by expression of Lgr5, they are actively cycling and give rise to cells that proliferate in the transiently amplifying compartment of the crypt. Cells move up from the TA compartment and differentiate in the
invaginations of the intestinal epithelium; the crypts of Lieberk��hn. The intestinal stem cells are characterized by expression of Lgr5, they are actively cycling and give rise to cells that proliferate in the transiently amplifying compartment of the crypt. Cells move up from the TA compartment and differentiate in the