These results are seen for both HH and HF individuals, suggesting that the reasons for the changed ultrastructure is primarily due to the high SF levels. In whole blood smears without thrombin, but with added sodium salicylate, RBCs do not have the pointed extensions. However, with added thrombin, most of the RBCs seem to be folded around the fibrin fibers, changing the typical discoid shape. Fibrin Benzoylaconine fibers were comparable to those of healthy individuals. Lower concentrations of the additives did stabilize the RBC as well as fibrin fiber morphology. Light microscopy of whole blood with these lower concentrations also shows the stabilizing of the RBCs. The same pattern was seen in the presence of sodium selenite, except that it seems as if the RBC kept their shape better in the presence of the sodium selenite and thrombin. Clioquinol showed anomalous results, where RBC with and without thrombin kept their pointed appearance and the fibrin fibers also coagulated into DMDs with very few individual fibers visible. The lower additive concentrations for all products, show a less prominent stabilizing effect when viewed with SEM. However, light microscopy of samples in the presence of the lower concentrations clearly show that most of the RBCs have returned to the discoid shape. This was noted for HH as well as wild type/wild type individuals with higher than the accepted healthy serum iron levels. Individuals with hemochromatosis �C as an ‘iron overload disease’�C are well known 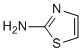 to have significantly more ��iron’in their bodies than do normal controls, and this is considered to contribute to the Tubeimoside-I attendant gross pathologies of this syndrome. Phlebotomy and iron chelation are thus two therapies in common use. Here we establish that accompaniments of this excessive iron in hemochromatosis whole blood are changes in both RBC morphology and in fibrin fibers, possibly due to hydroxyl radical formation or to the presence of excess iron itself. If hydroxyl radicals are involved, we suggest that they can cause non-enzymatic changes to fibrin in the presence of thrombin and a changed RBC ultrastructure, where the cells lose their discoid shape and are easily deformed when fibrin and DMDs are produced in the presence of thrombin. Ferric ions may also bind to the outer surface of the RBC directly. As expected, the classical iron chelator, desferal, showed a stabilizing effect on both fibrin fiber and RBC ultrastructure. Clioquinol is known to chelate and redistribute iron. However, in the current work, it showed the least potential to stabilize RBCs and fibrin. By contrast, salicylate and sodium selenite showed excellent stabilizing properties. Salicylate is also known to be a direct free radical scavenger and recently it has been shown that it affords protection against rotenone-induced oxidative stress and therefore has neuroprotective potential against OHN radical damage. In the current study, sodium selenite and sodium salicylate plausibly also inhibited the hydroxyl radicals produced by the increased iron present in hemochromatosis, but as mentioned above may well also have bound or chelated some of the free iron. The current research has shown that iron causes structural changes, but that selected additives cause a reverting of the structure; this suggests that the damage seen is indeed reversible. As discussed in the previous paragraphs, iron causes oxidative stress in cells. However, in the current manuscript we did not look at the specific markers that might cause oxidative damage, e.g. the presence of ROS in the RBCs. Some of the effects might be purely due to binding, e.g. via electrostatic effects.
to have significantly more ��iron’in their bodies than do normal controls, and this is considered to contribute to the Tubeimoside-I attendant gross pathologies of this syndrome. Phlebotomy and iron chelation are thus two therapies in common use. Here we establish that accompaniments of this excessive iron in hemochromatosis whole blood are changes in both RBC morphology and in fibrin fibers, possibly due to hydroxyl radical formation or to the presence of excess iron itself. If hydroxyl radicals are involved, we suggest that they can cause non-enzymatic changes to fibrin in the presence of thrombin and a changed RBC ultrastructure, where the cells lose their discoid shape and are easily deformed when fibrin and DMDs are produced in the presence of thrombin. Ferric ions may also bind to the outer surface of the RBC directly. As expected, the classical iron chelator, desferal, showed a stabilizing effect on both fibrin fiber and RBC ultrastructure. Clioquinol is known to chelate and redistribute iron. However, in the current work, it showed the least potential to stabilize RBCs and fibrin. By contrast, salicylate and sodium selenite showed excellent stabilizing properties. Salicylate is also known to be a direct free radical scavenger and recently it has been shown that it affords protection against rotenone-induced oxidative stress and therefore has neuroprotective potential against OHN radical damage. In the current study, sodium selenite and sodium salicylate plausibly also inhibited the hydroxyl radicals produced by the increased iron present in hemochromatosis, but as mentioned above may well also have bound or chelated some of the free iron. The current research has shown that iron causes structural changes, but that selected additives cause a reverting of the structure; this suggests that the damage seen is indeed reversible. As discussed in the previous paragraphs, iron causes oxidative stress in cells. However, in the current manuscript we did not look at the specific markers that might cause oxidative damage, e.g. the presence of ROS in the RBCs. Some of the effects might be purely due to binding, e.g. via electrostatic effects.
The typical net does not completely form in the manner seen with the higher desferal concentration
Leave a reply